The initial steps of HR involve processing the DNA ends of DSBs by exonucleases to generate 3’ single-stranded DNA (ssDNA) tails (Figure 1). Two recombinases, Rad51 and Dmc1, then form nucleoprotein filaments on ssDNA and search for DNA homologous sequences leading to strand invasion (D-loops in in vitro assay). Remarkably, proper function of the recombinases requires interactions with several accessory proteins. For example, Hop2 and Mnd1 are two proteins essential for normal HR progression via their interactions with Dmc1 and Rad51. It is currently accepted that strand invasion intermediates proceed by one of two distinct pathways illustrated in Figure 1. The HR dissociate, which leads to re-joining of the broken chromosome by synthesis-dependent strand annealing (SDSA) to generate non-crossovers (NCO), or alternatively, they proceed by double strand break repair (DSBR), generating crossovers (CO). Regulation of CO/NCO products is critical in meiosis. Suppression of excess CO is indispensable for preservation of genome integrity. Equally important, however, a minimum critical number of CO is required to ensure proper segregation of meiotic chromosomes.
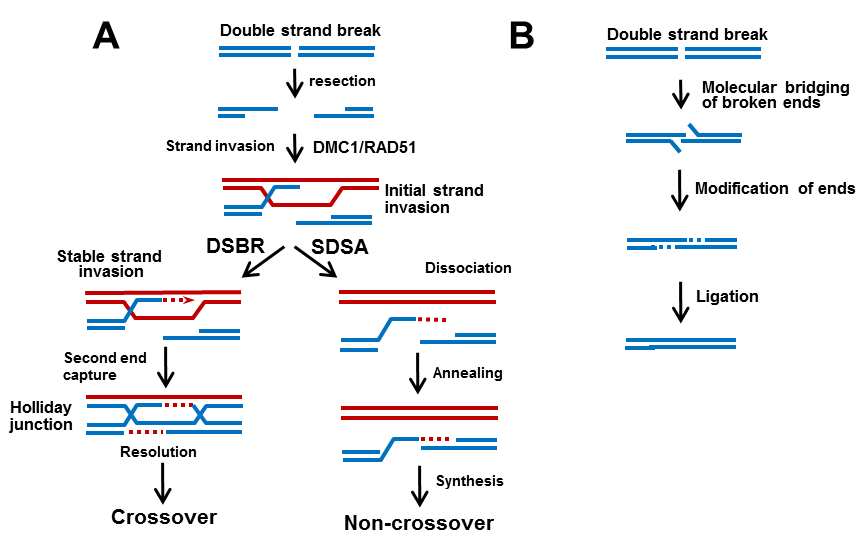
Figure 1. Homologous recombination and NHEJ DNA repair pathways. A. Red and blue represent DNA copies of two different homologous chromosomes. While SDSA only produce NCO, during DSBR the second DSB end is captured. After gap-filling DNA synthesis and ligation, it forms Holliday junction who resolution can result in the formation of most crossovers and a small fraction of noncrossovers. B. NHEJ rejoins broken ends.
The enzymatic machinery for recombination repair in mitosis is nearly identical to the machinery for DSBs repair during meiosis. In meiosis, HR also mediates the exchange of information between the maternal and paternal alleles, which generates genetic diversity in gametes. Importantly, at the end of meiotic prophase, HR serves a third critical function by providing a physical link that holds homologous chromosome pairs together. These linkages established by chiasmata, which are the cytological manifestation of the crossover product of HR, together with cohesin linkage between sister chromatids, ensure the orderly segregation of each chromosome in the pair to the opposite poles of the spindle resulting in gametes with the correct number of chromosomes (Figure 2A). Accurate DSB repair through recombination in meiosis is critical to avoid deleterious outcomes including miscarriages, infertility, and birth defects. Indeed, an estimated 10 to 30% of fertilized human eggs have the wrong number of chromosomes resulting in at least 5% of conceptions being aneuploid. Most of them abort before term making aneuploidy the leading known cause of pregnancy loss. Those who survive face devastating consequences, including developmental disabilities and mental retardation such as Down, Turner, and Klinefelter syndromes.